メニュー
果樹、野菜、花卉などの園芸植物の種苗は、種子繁殖と栄養繁殖(培養も含む)によって生産されます。種子繁殖は、受粉、花粉発芽、花粉管伸張、受精、胚の発達、種子形成といった一連の有性生殖の過程を踏みます。この過程は果実の結実・発育にも重要で、種子が形成されない多くの果実は発育を停止し、落果してしまいます。一方、栄養繁殖は、植物体の一部を挿木、接木、株分け・分球、培養する増殖法で、親植物のヘテロ性が高く、種子繁殖すると形質が分離してしまう多年草の草本や木本植物の繁殖に用いられています。園芸植物繁殖学では、園芸植物の生殖・繁殖のメカニズムを解明し, それを制御することで、品種育成、種苗生産や果実生産に貢献するための教育と研究に取り組でいます。現在の主な研究テーマには以下の通りです。
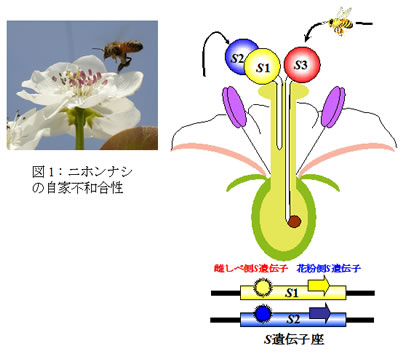
1つの花の中に雌しべと雄しべがある植物の繁殖様式は、自分の花粉(自家花粉)で受精・結実する自殖性と、自家花粉では受精・結実できず、他個体の花粉が受粉され、受精・結実する他殖性に大別されます。自家花粉の発芽および花粉管伸長を妨げ、自殖を抑制する現象は「自家不和合性」と呼ばれ、植物が近交弱勢を回避し、他殖を促進することで、遺伝的多様性を広げるために獲得した形質です。
ナシ属の自家不和合性、遺伝学的に配偶体的に機能する1遺伝子座のSハプロタイプ (S1、S2、S3、…、Sn) によって制御されます。
雌しべと花粉が同じ番号のSを持つ場合、不和合となり、花粉管伸長は停止します。異なる番号の持つ場合、和合になります(図1)。この雌しべと花粉間における自他認識反応は雌しべ側S遺伝子と花粉側S遺伝子産物の相互作用により起こります。分子生物学および細胞組織学的手法を用いて、自家不和合性のメカニズムの解明に取り組んでいます。

植物では、同一種内あるいは異種間の交雑によって生じた雑種が両親よりも優れた形質を示す『雑種強勢/ヘテローシス』という現象が知られています。現在、多くの野菜は、異なる性質をもつ両親系統間の交雑によって得られた雑種第一代 (F1: First Filial Generation)の種子を大量に採種し、品種とする一代雑種 (F1)品種です。F1品種には、「両親系統の優良形質を同時に付与できる。」や「形質が均一である。」といった特徴に加え、「雑種強勢により生育が旺盛で収量性が向上する。」といった特徴があります (図2)。よって、雑種強勢は、植物の収量増加に貢献した遺伝現象として知られており、F1品種による収量改善効果が著しいことは既に明白ですが、未だ、雑種強勢の分子機構は、ほとんど明らかになっていません。そこで、雑種強勢の分子機構の解明を目指して、ハクサイやキャベツといったアブラナ科野菜を中心に、分子生物学的な研究を進めています。
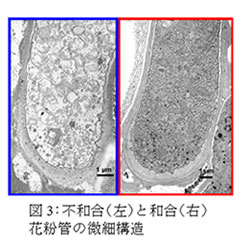
自家不和合性の雌しべと花粉間の自他認識反応はS遺伝子座上の雌しべ側S遺伝子であるS-RNaseと花粉側S遺伝子産物の相互作用により起こります。花粉管に取り込まれた自己S-RNaseは不和合花粉管のrRNAを分解し、花粉管内部の微細構造を崩壊させます(図3)。
一方、和合花粉管に取り込まれた非自己S-RNaseは花粉側S遺伝子産物により阻害・分解させると考えられてきました。
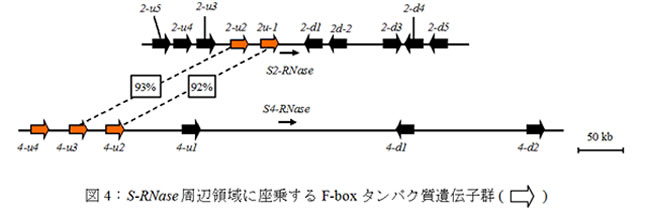
花粉側S遺伝子を探索するため、ナシのBACライブラリーを作成し、S-RNaseを起点とした染色体歩行によりS-RNase周辺領域BACコンティグを構築しました。塩基配列解析の結果、S-locus F-box Brothers (SFBB)と呼ばれるF-boxタンパク質遺伝子群が見つかりました (図4)。
F-boxタンパク質は結合した標的タンパク質をユビキチン化し、26Sプロテアソームによるタンパク質分解へと導きます。ハプロタイプ間で相同性(%)が高いSFBBは同一のS-RNaseを認識している可能性があります (図4)。
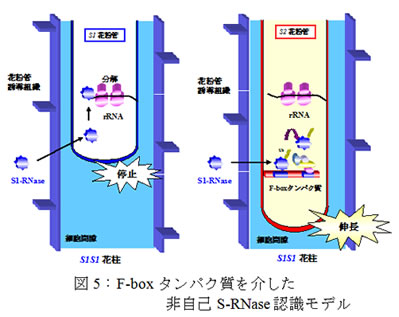
SFBB群は分担して非自己S-RNaseを認識(結合)してユビキチン化している可能性が考えられます (図5)。

雑種強勢の分子機構の解明を目指して分子生物学的な手法を用いて研究を進めています(図6)。具体的には、雑種強勢の形質と関連する遺伝子を見出す為に、F1品種とその両親系統の全遺伝子の転写産物を網羅的に比較するトランスクリプトーム解析を行っています。また、雑種強勢は量的な形質であることから、QTL (Quantitative trait locus)解析により雑種強勢関連領域及び遺伝子の特定を目指しています。これらの解析を行う際には、次世代シークエンサーを利用したRNA sequencing法等、新しい研究手法を積極的に導入して行っています。さらに、遺伝子の発現制御はその塩基配列によって決定されていることが分かっていますが、一方で、塩基配列が完全に同一でも遺伝子の発現が異なる場合があることも分かってきています。このような塩基配列に依存 しない遺伝子の発現制御は、エピジェネティックな発現制御とよばれています (藤本•石川 2010, 育種学研究12: 123-131)。エピジェネティックな制御が雑種強勢と関わる可能性が示唆されていることから、ゲノム全体のエピジェネティックな修飾状態を調べることで、エピジェネティックな修飾状態と雑種強勢の現象との関連性について調べています。




